
DNA breaks activate the DNA damage response and, if left unrepaired, trigger cellular senescence. Telomeres are specialized nucleoprotein structures that protect chromosome ends from persistent DNA damage response activation. Whether protection can be enhanced to counteract the age-dependent decline in telomere integrity is a challenging question. Telomeric repeat–containing RNA (TERRA), which is transcribed from telomeres, emerged as important player in telomere integrity. However, how human telomere transcription is regulated is still largely unknown. We identify nuclear respiratory factor 1 and peroxisome proliferator–activated receptor g coactivator 1a as regulators of human telomere transcription. In agreement with an upstream regulation of these factors by adenosine 5′-monophosphate (AMP)–activated protein kinase (AMPK), pharmacological activation of AMPK in cancer cell lines or in normal nonproliferating myotubes up-regulated TERRA, thereby linking metabolism to telomere fitness. Cycling endurance exercise, which is associated with AMPK activation, increased TERRA levels in skeletal muscle biopsies obtained from 10 healthy young volunteers. The data support the idea that exercise may protect against aging.
INTRODUCTION
Mammalian telomeres are specialized structures that protect eukaryotic chromosome ends from DNA repair reactions, owing to a lariat conformation and the presence of a six-subunit protein complex, called shelterin (1). In humans, telomeres shorten with age, and this interferes with tissue replenishment (2). However, recent studies acknowledged that nongenetic influences can affect telomere attrition and possibly delay aging-related diseases (2, 3). Human telomeres are transcribed from CpG island–containing subtelomeric promoters, located on average 1 kb upstream ofTTAGGGrepeats, into telomeric repeat–containing RNA (TERRA)molecules (4, 5). TERRA molecules comprise subtelomerespecific sequences at their 5′ end,followed by stretches ofUUAGGGrepeats proportional to telomere length (6), and remains partly associated with telomeres to play crucial protective functions (7). Additional regulatory functions of TERRA will likely emerge in the future. In cycling cells, TERRA levels decline from S phase to G2 before increasing again, ensuring renewal of TERRA pools at each cell division (6, 8, 9). How human telomere transcription is activated is largely unknown, but transcriptional regulators of telomeres include CTCF and ATRX chromatin remodelers, as well as Rad21 cohesin subunit (10–12). Understanding the regulatory mechanisms of human telomere transcription, not only in cycling cells but also in nondividing tissues, is likely to provide new clues for antiaging research. To gain insight into the regulatory mechanisms of human telomere transcription, we screened the available human subtelomeric sequences for the presence of predicted transcription factor binding sites using an in silico approach. This allowed us to identify nuclear respiratory factor 1 (NRF1) as a key regulator of human telomere transcription and revealed a new link between telomeres and metabolism.
RESULTS NRF1
binds human subtelomeres To identify new transcription factors for TERRA, we performed an in silico analysis of human subtelomeric sequences. The “low-confidence” data set (Fig. 1A and table S1) refers to subtelomeric sequences from the study by Stong et al. (13) that were either missing in the GRch38/hg38 assembly or lacking TTAGGG repeats at their 3′ ends. Sequences of p arms from acrocentric chromosomes are not available. On the basis of a previous study by Nergadze et al. (5), we identified putative transcription start sites (TSSs) on 18 of 27 and 4 of 14 subtelomeres from the high- and low-confidence data sets, respectively (Fig. 1A). MatInspector (14) revealed the presence of putative binding sites for NRF1 in 30 of 41 subtelomeric sequences. NRF1 is an important regulator of nucleus-encoded subunits of mitochondrial respiratory complexes, and its disruption leads to early mouse embryonic lethality associated with severe mitochondrial DNA depletion (15). NRF1 targets are coactivated by PGC-1a (peroxisome proliferator–activated receptor g coactivator 1a), a key regulator of energy metabolism that is activated by endurance exercise or caloric restriction and previously linked to protection against aging (16). This prompted us to further investigate the link between NRF1 and telomere transcription. We first set up chromatin immunoprecipitation (ChIP) experiments. Knowing that NRF1 binds DNA within CpG-rich sequences and is sensitive to methylation (17), we selected LB37 non–small cell lung carcinoma cell line with hypomethylated subtelomeric promoters and high TERRA expression levels (6) to assess NRF1 binding. Because we could not design primers suitable for quantitative polymerase chain reaction (qPCR) within the repetitive CpG-rich sequences of subtelomeric promoters, qPCRs amplified genomic loci located either directly upstream (21q, 10p-18p) or directly downstream (5p, Xq-Yq, 1q-2q-4q-5p-10q-13q-21q-22q, 15q) of CpG islands. In agreement with in silico analysis, NRF1 binding was detected on all subtelomeric sequences with predicted NRF1 binding sites that we tested [from about 5- to 50-fold enrichment over immunoglobulin G (IgG); Fig. 1B]. 7q subtelomere, which lacks predicted TSS and displays only one putative NRF1 binding site, showed enrichment over IgG of only 1.9-fold (seefig. S1A for primer position), in accordance with the extremely low transcriptional activity of that subtelomere (Fig. 1, A to C, andfig. S1, A to D). As expected, NRF1 did not bind to 10p-18p subtelomeric loci located about 1.5 kb upstream of telomeres (Fig. 1B, 10p-18p distal; 1.6-fold over IgG). Furthermore, we could show that the ability of NRF1 to bind 15q subtelomere correlates with 15q TERRA levels in LB37 and Huh-7 hepatocarcinoma cell lines and inversely correlates with the distance from CpG island (Fig. 1, D and E). Together, the above data suggested that NRF1 may play a role in human telomere transcription. Cycling endurance exercise up-regulates TERRA levels in human skeletal muscle Having shown that NRF1 binds human subtelomeres, we next tested the hypothesis that endurance exercise, a well-established inducer of NRF1 target genes, may affect telomere transcription. Briefly, adenosine 5′-diphosphate (ADP)/adenosine 5′-triphosphate (ATP) ratio increases during exercise, leading to adenosine 5′-monophosphate (AMP)–activated protein kinase (AMPK) activation. In turn, activated AMPK phosphorylates PGC-1a to promote its nuclear accumulation through sirtuin 1–dependent deacetylation. Once in the nucleus, PGC-1a acts as transcriptional coactivator for various transcription factors, including NRF1 (16, 18). To investigate the impact of exercise on telomere transcription in skeletal muscles, we submitted 10 healthy young volunteers to a cycling endurance exercise for 45 min. To obtain various levels of AMPK activation, subjects were submitted to either low-intensity (50% VO2 peak) or high-intensity (75% VO2 peak) exercise, associated with, respectively, low or high AMPK activation levels. Three muscle biopsies were taken according to a standardized protocol either before (B1), directly after (B2), or 2.5 hours after (B3) exercise (Fig. 2A). Blood lactate was immediately measured at the end of the exercise to evaluate individual responses (Fig. 2B). Fitting with post-exercise blood lactate levels, phosphorylation of acetyl–coenzyme A carboxylase (ACC) in muscle biopsies,a bona fide marker of AMPK activation, increased with response intensity
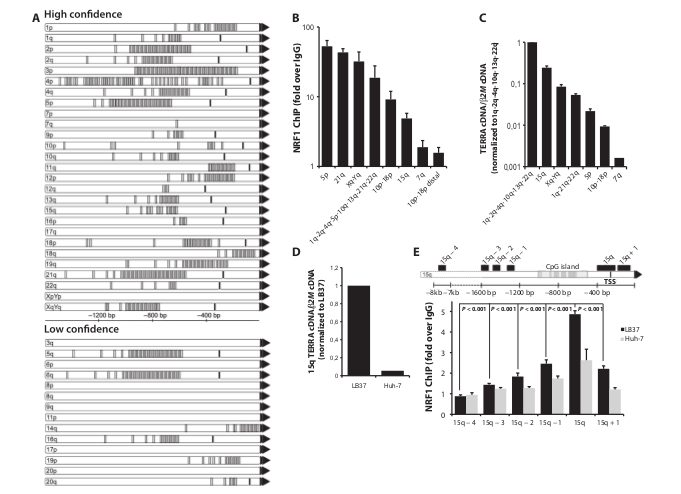
Fig. 1. NRF1 binds human subtelomeric promoters. (A) Predicted NRF1 binding sites on human subtelomeres (gray bars). Black bars indicate putative TSS based on the study by Nergadze et al. (5). Black triangles indicate telomeres. (B) NRF1 binding at subtelomeres of LB37 cells. Graph shows fold enrichment over IgG. Error bars indicate SD (n = 3). (C) qRT-PCR analysis of TERRA in LB37 cells for the indicated chromosome ends. TERRA cDNA levels were first normalized to b2M cDNA and then to the relative expression level of 1q-2q-4q-10q-13q-22q TERRA. Error bars indicate SD (three independent RNA extractions). (D) Relative 15q TERRA expression in LB37 and Huh-7 cell lines (normalized first to b2M cDNA and then to LB37). (E) NRF1 binding assessed by ChIP on six loci spread onto 15q subtelomere in LB37 and Huh-7 cell lines. Graph shows fold enrichment over IgG. Error bars indicate SD (n = 3).
(Fig. 2,C to E).Accordingly, nuclear translocation of PGC-1a, a marker of its activation, was detected in B2 and B3 samples (Fig. 2F). PGC-1a nuclear translocation increased with blood lactate concentration and was up-regulated by a factor of 2 in B3 biopsy from S5 (high lactate) compared to S12 (low lactate) (Fig. 2F). In agreement with activated PGC-1a up-regulating its own transcription (18), PGC-1a mRNA levels were increased by up to 37-fold in B3 samples (Fig. 2G). The lack of induction of PGC-1a mRNA in B2 perfectly fits with previous observations in human muscle, where up-regulation of PGC-1a mRNA was very weak immediately at the end of exercise but peaked within 2 hours after exercise bout (19). Strikingly, quantitative reverse transcription PCR (qRT-PCR) against distinct TERRA 5′ ends revealed up-regulation in 50 to 90% of B2 samples and in 80 to 100% of B3 samples, depending on the chromosome end tested (Fig. 2H). Compared to matching B1, TERRA levels in B3 reached an average of 186 and 131% in the high- and low-intensity exercise group, respectively (Fig. 2I). The different induction timing observed for TERRA (already in B2) and PGC-1a (in B3) transcription may result from distinct mechanisms of PGC-1a coactivation. As a transcriptional coactivator, PGC-1a interacts with multiple and various DNA binding factors, the nature of which depends on the target gene. For PGC-1a transcription, NRF1 is not involved (20). Our data fit with the observation that, in response to exercise, NRF1-dependent mitochondrial biogenesis occurs before the up-regulation of PGC-1a levels in rat muscles (21). Plotting TERRA induction in B3 against post-exercise blood lactate concentration revealed a significant correlation (P < 0.05) (Fig. 2J). Because blood lactate concentrations correlated with AMPK activity in muscle tissues (P < 0.005) (Fig. 2E), these data suggest that the kinase regulates telomere transcription. Together with our demonstration that most telomeres from muscle cells are probably covered with TERRA (Fig. 2K and fig. S2), this finding suggests that exercise provides a means to renew TERRA pools and protect telomeres in muscle.
NRF1 and AMPK/PGC-1a axis promote human telomere transcription
The endurance exercise experiment suggested that telomere transcription is regulated by the AMPK pathway. However, although NRF1 is expressed in skeletal muscles (fig. S3), our in vivo experiment did not allow us to test whether the transcription factor is implicated in telomere transcription. To further investigate this, and to gain more insight into AMPK-dependent regulation of TERRA, we used the Huh-7 cell line that responds to phenformin, a biguanide drug that, like metformin, activates AMPK by increasing cellular AMP/ATP ratio (22). First, we showed that either NRF1 knockdown or overexpression of a dominant negative form that lacks the C-terminal transactivation domain (DC NRF1) (23) reduces endogenous TERRA levels by 25 to 45% (Fig. 3, A and B), supporting a role for NRF1 in basal transcription of Huh-7 telomeres. Accordingly, overexpression of NRF1 stimulated luciferase activity driven by a portion of 10q promoter (10) containing NRF1 binding sites, whereas DC NRF1 had opposite effects (Fig. 3C). Second, when Huh-7 cells were treated with phenformin, ACC phosphorylation was increased, PGC-1a accumulated in the nucleus, and TERRA levels, measured from various chromosome ends, reached 185 to 400% of the expression detected in untreated cells (Fig. 3, D to F). PGC-1a transcriptional induction also occurred, whereas no noticeable change was observed for the three control genes: hTR noncoding telomerase RNA subunit, TRF2 shelterin gene, or b2M which was used to normalize complementary DNA (cDNA) values (Fig. 3F and fig. S4, A and B). Here, too, PGC-1a mRNA up-regulation occurred later than TERRA induction (fig. S4B). In agreement with AMPK activation acting at the level of telomere transcription, phenformin treatment induced 10q-luciferase activity by a factor of 2; induction was further exacerbated by overexpression of wild-type NRF1 (3.3-fold) but mostly lost upon DC NRF1 overexpression (Fig. 3G). Mutation of NRF1 binding sites in the 10q promoter-luciferase construct (fig. S5) reduced basal activity of the promoter by 35% (mut3) and 70% (mut4), respectively, and lowered its activation by either overexpressed NRF1 or phenformin treatment (Fig. 3H). Next, to probe more directly for PGC-1a involvement in TERRA transcription, we transduced Huh-7 cells with adenoviral particles containing either mouse PGC-1a (mPGC-1a) coding sequence or GFP cDNA as control (fig. S6A). Luciferase activity driven by 10q promoter was up-regulated by a factor of 4.7 upon mPGC-1a overexpression and of 13.8 when cells were simultaneously overexpressing wild-type, but not DC, NRF1 (Fig. 3I). Accordingly, mPGC-1a overexpression up-regulated endogenous TERRA levels by factors of 1.4 to 1.6 (Fig. 3J). Although modest, the induction was significant and only twofold less than the induction of hCYTC gene, a well-established PGC-1a target gene (fig. S6B) (24). However, we could not detect any strong enrichment of NRF1 binding onto TERRA promoter in the presence of overexpressed PGC-1a, suggesting that posttranslational modifications of prebound NRF1 (15), rather than a massive recruitment of the transcription factor, may be involved in telomere transcription activation. This observation fits with the report that PGC-1a overexpression fails to increase NRF1 occupancy on promoters of TFB1M and TFB2M genes encoding two mitochondrial transcription specificity factors (24). Together, the above data demonstrated a pivotal role of NRF1 and the AMPK/PGC-1a axis in promoting human telomere transcription. Given the various protective functions that TERRA plays at telomeres (7), we next tested whether NRF1 down-regulation may impair telomere integrity by analyzing the formation of telomere dysfunction– induced foci (TIF) in Huh-7 cells. Accordingly, small interfering RNA (siRNA)–mediated loss of NRF1 increased the occurrence of colocalization events between telomeres and 53BP1, a marker of DNA damage, from an average of 1.8 per nucleus in siLuci-treated cells to 3.8 per nucleus upon siNRF1 treatment (P < 0.001) (Fig. 3K). These data support an important role of NRF1 in maintaining human telomere integrity, a function that likely involves its ability to promote telomere transcription. However, the number of 53BP1 foci induced by NRF1 depletion was not very high, and foci were quite large. In light of the demonstration that chromosomal lesions induced by incomplete DNA replication are sequestered in 53BP1-enriched nuclear compartments of G1 cells to protect them from erosion (25), it is tempting to speculate that NRF1 may participate in telomere replication. One possibility would be that, on the basis of evidences obtained from budding and fission yeast (26, 27), NRF1-dependent TERRA production may help in recruiting telomerase at chromosome ends to complete telomere synthesis. Alternatively, the transcription process per se may possibly help in resolving topological barriers at telomeres to promote replication fork progression.
AMPK activation induces NRF1-dependent TERRA up-regulation in human myotubes
Because the above experiments were performed in cycling cancer cells, we next tested the relevance of our observations in the physiological context of healthy nondividing muscle cells. To this end, we differentiated human deltoid-derived myoblasts into myotubes (fig. S7). To our knowledge, the presence of TERRA at telomeres of these cells has not been checked before. Combined RNA/DNA–fluorescence in
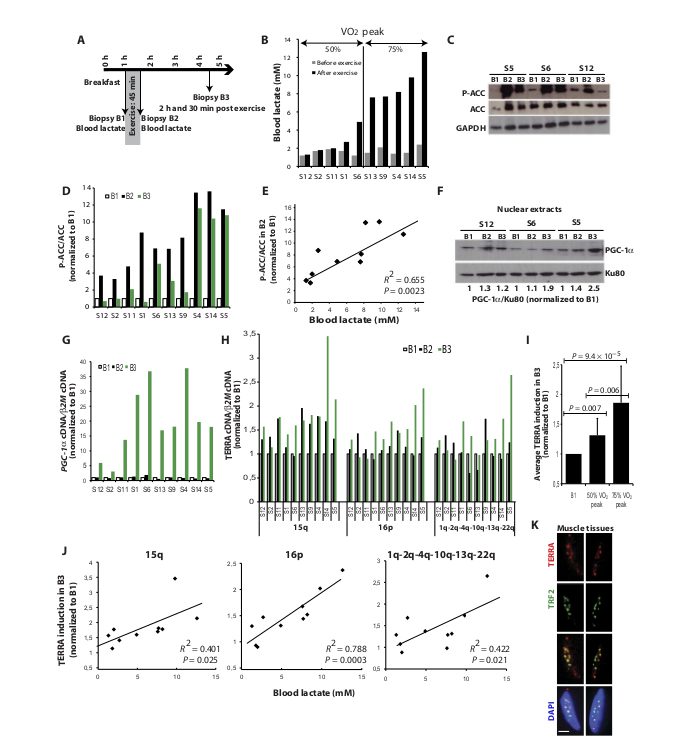
Fig. 2. Endurance exercise up-regulates TERRA in human muscle. (A) Design of the in vivo experiment. (B) Blood lactate concentrations (mM) before (gray) and at the end (black) of exercise in subjects (S) (50% VO2 peak, low-intensity exercise; 75% VO2 peak, high-intensity exercise). (C) Representative Western blots of ACC phosphorylation (P-ACC) in B1, B2, and B3 biopsies from S5 (high lactate), S6 (medium lactate), and S12 (low lactate). Total ACC was used to evaluate the P-ACC/ACC ratio, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a loading control. (D) P-ACC/ACC ratios in B2 and B3 normalized to B1 for all subjects. (E) P-ACC/ACC ratios plotted against blood lactate concentration after exercise. (F) PGC-1a abundance in nuclear fractions from S12, S6, and S5 biopsies normalized to Ku80 and to matching B1. (G) PGC-1a cDNA normalized to b2M cDNA and to matching B1. (H) qRT-PCR analysis of TERRA (15q, 16p, and 1q-2q-4q-10q-13q-22q) in B2 and B3, normalized to b2M cDNA and to matching B1. (I) Average induction of TERRA (all three qRT-PCRs pooled) in B3 compared to matching B1 for 50% VO2 peak group (n = 5) and 75% VO2 peak group (n = 5). Error bars indicate SD. (J) TERRA fold induction in B3 plotted against blood lactate concentration after exercise. (K) TERRA-FISH (red) and TRF2 detection (green) in muscle biopsies. Blue, 4′,6-diamidino-2- phenylindole (DAPI). Scale bar, 5 mm.
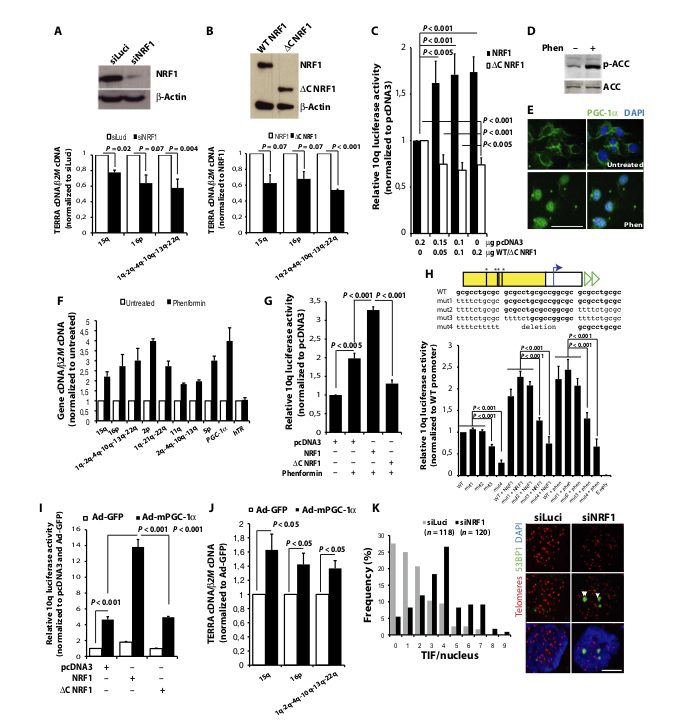
Fig. 3. NRF1 and AMPK/PGC-1a axis promote human telomere transcription. (A) NRF1 protein in siLuci- and siNRF1-treated Huh-7 cells; TERRA cDNA normalized to b2M cDNA and siLuci (n = 3). (B) NRF1/DC NRF1 overexpression in Huh-7 cells; TERRA cDNA normalized to b2M cDNA and to NRF1-overexpressing cells (n = 3). (C) Transcriptional activity of 10q promoter in Huh-7 cells overexpressing increasing amounts of NRF1/DC NRF1 (n = 3). (D and E) Phenformin treatment in Huh-7: ACC phosphorylation and PGC-1a nuclear translocation. Scale bar, 100 mm. (F) qRT-PCR analyses of TERRA, PGC-1a, and hTR upon phenformin treatment normalized to b2M cDNA and to untreated cells (n = 3). (G) Transcriptional activity of 10q promoter in phenformin-treated Huh-7 cells overexpressing NRF1 or DC NRF1 normalized to pcDNA3-transfected control cells (n = 5). (H) Impact of NRF1 binding site mutation on 10q transcriptional activity. Data were normalized to untreated cells transfected with wild-type (WT) 10q promoter and pcDNA3 (n = 4). Empty: No promoter. (I) 10q transcriptional activity upon mPGC-1a overexpression, in combination with NRF1/DC NRF1. Data were normalized to pcDNA3-transfected/adenovirus–green fluorescent protein (Ad-GFP)–transduced cells (n = 6). (J) TERRA cDNA in mPGC-1a–overexpressing Huh-7 normalized to b2M cDNA and Ad-GFP control cells (n = 5). (A to J) Error bars indicate SD. (K) TIF formation upon NRF1 knockdown in Huh-7 cells. Telomeres were detected by FISH (red) and DNA damage with 53BP1 antibody (green). Data are representative of three independent transfections. Scale bar, 5 mm
situ hybridization (FISH) experiments in myotubes revealed an average of 36 telomeric signals and 38 TERRA foci per nucleus (Fig. 4, A and B, and fig. S8). Telomeric signal abundance agrees with a previous report that telomeres are clustered in human cells (28). An average of 35 TERRA foci colocalized with telomeres, suggesting that, although not all telomeres are transcribed, about 97% of them may be covered by TERRA if RNA molecules were provided in trans. Conversely, and for still unknown reasons, two to three TERRA foci per nucleus were not associated with telomeres. AMPK pathway activation in primary myotubes treated with either phenformin, metformin, or AICAR (5-aminoimidazole-4-carboxamide ribonucleotide), another well-known AMPK activator (18, 22), led to ACC phosphorylation (Fig. 4C) and was associated with consistent and significant up-regulation of TERRA levels by factors of 1.6 to 2.2 (P < 0.001; Fig. 4D). To assess NRF1 involvement in AMPK-induced TERRA levels, we transfected myoblasts on day 1 of differentiation with siNRF1, before treatment with phenformin at day 4 and harvesting at day 5. The increase in TERRA expression upon phenformin treatment was lost when cells were transfected with siNRF1 but not with nonrelevant siRNA (siLuci) (Fig. 4, E and F). The absence of siNRF1 impact on basal TERRA levels may be due to the fact that myotubes are noncycling cells and TERRA turnover may thus be different from cells in which degradation is linked to cell cycle progression (8). In this view, it is expected that longer incubation times with siNRF1 may be required to detect any impact on basal TERRA levels. However, this extended treatment would have a negative impact on cell integrity. Together, these results show that AMPK activation induces NRF1-dependent increase in TERRA levels in noncycling human myotubes.
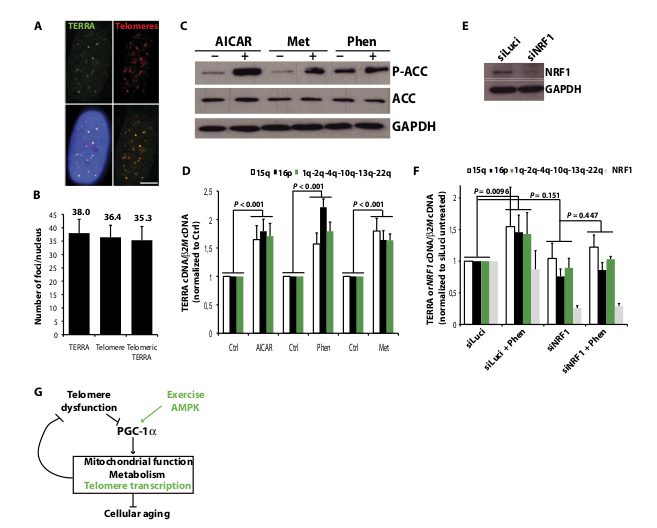
Fig. 4. AMPK activation in human myotubes induces NRF1-dependent increase in TERRA levels. (A) TERRA-FISH (green) combined with telomeric DNA FISH (red) in myotubes. Blue, DAPI. Scale bar, 5 mm. (B) Quantification of (A) on 25 nuclei. (C) ACC phosphorylation in myotubes treated with either AICAR, metformin, or phenformin. (D) qRT-PCR analysis of TERRA levels in treated myotubes normalized to b2M cDNA and to untreated cells. Error bars indicate SD (n = 4). (E) Western blot analysis of NRF1 knockdown in myotubes. (F) qRT-PCR analysis of TERRA in siNRF1-treated myotubes and upon phenformin treatment. Values were normalized to b2M cDNA and to siLuci-treated cells without phenformin. Error bars indicate SD (n = 3). (G) Unified theory of aging (34) revisited with data from this study (green). See text for details.
DISCUSSION
The association between physical activity and telomere length is still an open question. Although endurance training may provide protective effects on telomere length and attenuate biological aging (29), acute exposure to ultradistance endurance trail races was reported to shorten telomeres (30) as a possible consequence of excessive reactive oxygen species (ROS) production (31). Because we found that telomere transcription is activated by NRF1 antioxidant factor, it is tempting to speculate that TERRA up-regulation may be part of the antioxidant response that skeletal muscles set up to counteract exercise-induced ROS (32). In this view, we hypothesize that, because of their high content in guanine residues, UUAGGG repeats may be highly prone to oxidation and telomere-bound TERRA may possibly shield TTAGGG telomeric repeats from ROS. TERRA oxidation may also occur as a daily consequence of cellular respiration, and exercise would thus help in renewing TERRA pools by eliminating oxidized molecules, thereby ensuring maintenance of TERRA-protective functions. It would be interesting to test that hypothesis in the future. Alternatively, the telomere transcription process per se may positively affect telomeres, possibly through chromatin remodeling or telomeric loop formation. TERRA also recently emerged as a telomerase regulator, but it is still unclear whether telomeric RNA species act as positive or negative regulators of telomerase (7). The demonstration that short telomeres of budding yeast cells display higher transcriptional activity to promote telomerase recruitment (26), together with similar data recently obtained from fission yeast (27), favors the positive regulator hypothesis. Although our data show that AMPK activation up-regulates TERRA regardless of telomerase expression status (Huh-7 cells express telomerase, whereas myotubes do not), we propose that exercise may also positively affect telomeres by promoting telomerase recruitment at telomeres of somatic stem cells that express the enzyme. Our study also provided a new link between telomeres and PGC-1a. First connections between telomeres, mitochondria, and PGC-1a were established by Sahin and colleagues (33), who reported that telomere dysfunction compromises mitochondrial and metabolic functions in mice through transcriptional repression of PGC-1a/b. Hence, in the context of the unified theory of aging involving PGC-1 as a key regulator (34), we propose that AMPK-activated transcription of telomeres may be part of a compensatory mechanism that aims to prevent telomere dysfunction–induced decline of postmitotic tissues and stem cells (Fig. 4G). These observations agree with the report that exerciseinduced up-regulation of PGC-1a reverts sarcopenia and, more generally, rescues progeroid aging in mitochondrial DNA mutator mice (35). Finally, our study agrees with the recent classification of dietary restriction, yet another AMPK/PGC-1a activation mechanism, with exercise, metformin, and sirtuin activators as possible interventions with translational potential that may delay aging (2, 3).
MATERIALS AND METHODS
In silico analysis of human subtelomeric promoters
Subtelomeric sequences obtained from the University of California, Santa Cruz genome browser using the December 2013 human reference sequence assembly (GRCh38/hg38)were assigned to the “high-confidence” data set. Missing subtelomeric sequences or sequences that did not have TTAGGG telomeric regions at the 3′ ends in GRCh38/hg38 assembly were obtained from the study by Stong et al. (13) and were classified as low confidence. Sequences of p armsfrom acrocentric 13, 14, 15, 21, and 22 chromosomes are missing. For promoter identification, 1.6 kb of upstream regions from final TTAGGGwas analyzed using the Genomatix software with PromoterInspector for the identification of putative RNA polymerase II promoter regions and MatInspector for the identification of putative transcription factor binding sites. Predicted TSSs were based on the study by Nergadze et al. (5).
Cell culture, transfections, and treatments
Huh-7 hepatocarcinoma cell line (American Type Culture Collection) was grown in Dulbecco’s modified Eagle’s medium (DMEM) (Lonza). LB37 non–small cell lung carcinoma cell line (6) was grown in Iscove’s modified Dulbecco’s medium (Lonza). Culture media were supplemented with 10% fetal bovine serum (FBS) (Gibco), 1% glutamine (Gibco), and 1% penicillin/streptomycin (Lonza). Transfections with siRNAs were performed as previously described (6) using Lipofectamine 2000 (Invitrogen) and either siNRF1 (table S2) or siLuci (6) (Eurogentec). For plasmid transfections, we used polyethylenimine (PEI) reagent (Polysciences, catalog no. 23966-2). Human myoblasts, provided by G. Carnac (INSERM U1046, University of Montpellier, France), were isolated from deltoid muscle biopsy and purified as previously described (36). Myoblasts were plated at a density of 1 × 104 cells/cm2 in DMEM supplemented with 20% FBS and 0.5% Ultroser G (Pall BioSepra). When cultures reached 80% of confluence, myogenic differentiation was induced by changing the medium to DMEM/2% FBS. Myotubes were obtained after 5 days. Cell treatments were performed as follows: 1 mM phenformin (Sigma-Aldrich) for 16 hours, 1 mM metformin (Sigma-Aldrich) for 8 hours, or 1 mM AICAR (Sigma-Aldrich) for 4 hours. For NRF1 knockdown in myotubes, myoblasts were transfected with siRNAs during the first day of differentiation using Lipofectamine RNAiMAX (Invitrogen), and cells were treated with phenformin on the fourth day of differentiation before harvesting 16 hours later. Ad-PGC-1a and Ad-GFP constructs were provided by B. Viollet and M. Foretz (Institut Cochin, Paris), and adenoviral particles were prepared using the human embryonic kidney 293 (HEK293) cell line according to standard procedures (37).
Human study
Ten healthy and moderately active young men [age, 20.0 ± 0.6 years; body mass index, 21.7 ± 0.7 kg m−2 ; peak oxygen consumption (VO2 peak), 47.2 ± 2.9 ml min−1 kg−1 ; maximal power output (Wmax), 226.9 ± 5.5 W] volunteered to participate in this study, which was approved by the local ethics committee (Université catholique de Louvain) and was in conformity with the Helsinki Declaration (www.wma.net/en/ 30publications/10policies/b3/index.html). Subjects were asked to refrain from vigorous physical activity for 2 days and to abstain from alcohol consumption the day before the experiments. A medical checkup was done to detect any contraindications for maximal exercise testing, and written consent was obtained from all subjects after all potential risks of the study were explained. At least 1 week before the experimental session, a maximal incremental exercise test was performed on a cycle ergometer (Cyclus2, RBM electronics) to determine VO2 peak and Wmax. The starting load was 70 W, incremented by 40 W every 3 min, until exhaustion. Heart rate (Polar) and respiratory exchanges (Medisoft) were continuously monitored. Participants were randomly divided into two groups: a low-intensity exercise group (50% VO2 peak) and a high-intensity exercise group (75% VO2 peak). On the day of the experimental session, subjects arrived after an overnight fast and received a standardized breakfast (715 kcal; 74% carbohydrate, 17% lipid, and 9% protein). One hour after breakfast, a first biopsy sample, with the needle pointing proximally, was taken from the midportion of the vastus lateralis muscle from the right leg under local anesthesia (1 ml of Xylocaine 2%; AstraZeneca). Samples were immediately frozen in liquid nitrogen and stored at −80°C before further analysis, except for ∼15 mg samples, which were embedded in the Tissue-Tek O.C.T. Compound (Sakura) for immunofluorescence (IF) staining and RNA-FISH experiments. A capillary blood sample (5 ml) was collected from the earlobe to determine the blood lactate concentration (Lactate Pro) before cycling exercise for 45 min at either 50 or 75% of VO2 peak. At the end of the exercise, the blood lactate concentration was measured again and a second biopsy was immediately taken in the same incision as the first one, with the needle pointing distally. Pointing the needle in the opposite direction was intended to reduce potential activation of inflammatory signaling pathways, as recommended (38). At 2.5 hours after the end of the exercise bout, a third biopsy was taken in the left leg.
Chromatin immunoprecipitation
ChIP experiments were performed as previously described (39) using NRF1 (table S3) or IgG control antibodies and OneDayChIP kit (Diagenode, catalog no. kch-504-100). Primers for PCR amplifications are described in table S2.
RNA extraction and qRT-PCR
RNA extraction, cDNA synthesis, and qPCR were performed as previously described (6) using KAPA SYBR FAST (KAPA Biosystems, catalog no. KK4608), and primers were described in table S2. For human muscle biopsies,RNAwas extractedfrom25 to 50mg of the sample homogenizedin1-ml TriPure reagent (Roche) using a T10 basic ULTRA-TURRAX (IKA).
Cellular fractionation from human muscle biopsies
About 20 mg of frozen muscle biopsies was homogenized with a Polytron mixer in 200 ml of ice-cold suspension buffer (20 mM Hepes, 5 mM NaF, 1 mM Na2MoO4, and 0.1 mM EDTA; protease inhibitor cocktail) (SigmaAldrich) supplemented with 0.5% NP-40 (Fluka). The homogenate was incubated on ice for 5 min and centrifuged at 10,000g for 30 s at 4°C. The supernatant was kept as cytosolic fraction. The pellet was then resuspended into 100 ml of suspension buffer containing 20% glycerol, 1 mM Na3VO4, 10 mM p-nitrophenyl phosphate, 10 mM b-glycerophosphate, and 5 mM NaF (Sigma-Aldrich) before 1:1 dilution with the same buffer containing 0.8 M NaCl. Samples were rotated for 30 min at 4°C before centrifugation at 10,000g for 10 min at 4°C, and supernatant was kept as nuclear fraction.
Western blotting
Total cell extracts were prepared with radioimmunoprecipitation assay buffer [150 mM NaCl, 50 mM tris-HCl (pH 8.0), 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS, 1 mM EDTA]. Western blotting was performed according to standard procedures using antibodies described in table S3. Revelation was performed with SuperSignal West Pico Chemiluminescent Substrate (Thermo Fisher Scientific, catalog no. 34080). Signal quantification was done using ImageJ software (National Institutes of Health).
IF, DNA-FISH, and RNA-FISH
IF was performed as previously described (39) using antibodies described in table S3. For TIF detection, IF was first performed using anti-53BP1 antibody (table S3). Then, cells were fixed again with 3.7% paraformaldehyde (PFA) (VWR)/phosphate-buffered saline (PBS) for 2 min at room temperature, washed with PBS, and treated with ribonuclease (RNase) A (100 mg/ml; Sigma-Aldrich) for 1 hour at 37°C before another incubation with permeabilization buffer for 10 min at room temperature. Cells were briefly washed with PBS, refixed with 3.7% PFA/PBS, and washed with PBS before successive baths in 70, 80, 90, and 100% ethanol for 2 min each. Hybridization with 50 nM TeloG Exiqon LNA red probe (TAMRA)GGGTtAGGGttAGgGTTAGGGttAGGGttAGGGtTA (TAMRA) (small letters indicate LNA-modified bases) was next performed in 50% deionized formamide (Millipore)/2× SSC in 1× blocking reagent (Roche) for 2 hours at room temperature after denaturation for 3 min at 85°C. Samples were dried out and mounted with DAPI (Sigma-Aldrich) after two washes with 50% formamide/2× SSC, 20 mM tris (pH 7.4) for 15 min at room temperature, three washes with 50 mM tris (pH 7.4), 150 mM NaCl, 0.05% Tween 20 (SigmaAldrich) for 5 min at room temperature and dehydration with ethanol bath series. RNA/DNA-FISH on primary myotubes differentiated onto coverslips was adapted from Basu et al. (40). Briefly, following TERRAFISH performed as previously described (6) with TeloC green Exiqon LNA probe (FAM)CCCTAaCcCTaaCcCTAACCCTaaCCCTaaCCCTaA(FAM) (small letters indicate LNA-modified bases), cells were fixed with 4% PFA (VWR) for 15 min at room temperature and rinsed once with PBS. Samples were next denatured at 80°C for 10 min in 70% deionized formamide (Millipore)/2× SSC before successive baths in 70, 85, and 100% ethanol. Denatured TeloG Exiqon LNA red probe (400 nM) was then added into the hybridization buffer [50% deionized formamide, 10% dextran sulfate (Pharmacia Biotech), 2× SSC, 0.2% bovine serum albumin (Sigma-Aldrich), 20 mM ribonucleoside-vanadyl complex (New England Biolabs)], and coverslips were incubated overnight at 37°C in a dark and humid environment. After three washes with 0.1× SSC at 37°C and one wash with 2× SSC, dehydration with ethanol bath series was performed again, and coverslips were dried out and mounted with DAPI (Sigma-Aldrich). TERRA-FISH combined with IF in muscle biopsies is described in Supplementary Methods. Images were acquired with a confocal microscope (Cell Observer Spinning Disc, Zeiss) with 100× objective and analyzed using the ImageJ software (National Institutes of Health).
Luciferase activity measurements
pGL3-10q construct containing the 10q subtelomeric promoter cloned upstream of Firefly luciferase gene (10) was provided by P. Lieberman (The Wistar Institute, Philadelphia). Luciferase activity normalization was obtained with pGL4.75(hRluc/CMV) vector containing the Renilla luciferase gene under the constitutive cytomegalovirus (CMV) promoter (Promega, catalog no. E6931). Mutation of NRF1 binding sites was performed using the QuikChange Multi Site-Directed Mutagenesis Kit (Agilent Technologies, catalog no. 200515) according to the manufacturer’s protocol, using primers described in table S2. Mutated DNA sequences are provided in fig. S5. For luciferase activity measurements, 8 × 104 Huh-7 cells per well were seeded into 12-well culture plates for 24 hours before transfection with PEI reagent (Polysciences, catalog no. 23966-2). Cells were transfected with a total of 0.5 mg DNA comprising 0.3 mg pGL3-10q, 0.005 mg pGL4.75, and 0.2 mg of either pcDNA3.1 empty plasmid, pcDNA3.1-Flag-NRF1, or pcDNA3.1-Flag-DC-NRF1 constructs (23) provided by K. Kohno (University of Occupational and Environmental Health Japan, Fukuoka). Cells were collected 24 hours after transfection, and relative luciferase activity was measured using the Dual-Luciferase Reporter Assay System (Promega, catalog no. E1910) on a GloMax 96 Microplate Luminometer (Promega). When stated, cells were treated with 1 mM phenformin (Sigma-Aldrich) for 16 hours.
Statistical analyses
Student’s t tests (Microsoft Excel) were applied to compare differences between means. For linear regressions, Pearson’s correlation coefficients and significance of R2 values were calculated using tools available at http://vassarstats.net/.
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/ content/full/2/7/e1600031/DC1 Supplementary Methods fig. S1. Analysis of 7q subtelomere. fig. S2. Control for TERRA-FISH in muscle biopsies. fig. S3. NRF1 is expressed in muscle biopsies. fig. S4. Time course of phenformin treatment in Huh-7 cells. fig. S5. Mutation of NRF1 binding sites in pGL3-10q construct. fig. S6. mPGC-1a overexpression in Huh-7 cells. fig. S7. Differentiated human myotubes. fig. S8. RNase control for TERRA-FISH in myotubes. table S1. Overview of available human subtelomeric sequences. table S2. Primers used in this study. table S3. Antibodies used in this study. References (41–45)
REFERENCES AND NOTES
1. N. Arnoult, J. Karlseder, Complex interactions between the DNA-damage response and mammalian telomeres. Nat. Struct. Mol. Biol. 22, 859–866 (2015).
2. E. H. Blackburn, E. S. Epel, J. Lin, Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 350, 1193–1198 (2015).
3. M. Kaeberlein, P. S. Rabinovitch, G. M. Martin, Healthy aging: The ultimate preventative medicine. Science 350, 1191–1193 (2015).
4. C. M. Azzalin, P. Reichenbach, L. Khoriauli, E. Giulotto, J. Lingner, Telomeric repeat– containing RNA and RNA surveillance factors at mammalian chromosome ends. Science 318, 798–801 (2007).
5. S. G. Nergadze, B. O. Farnung, H. Wischnewski, L. Khoriauli, V. Vitelli, R. Chawla, E. Giulotto, C. M. Azzalin, CpG-island promoters drive transcription of human telomeres. RNA 15, 2186–2194 (2009).
6. N. Arnoult, A. Van Beneden, A. Decottignies, Telomere length regulates TERRA levels through increased trimethylation of telomeric H3K9 and HP1a. Nat. Struct. Mol. Biol. 19, 948–956 (2012).
7. K. Rippe, B. Luke, TERRA and the state of the telomere. Nat. Struct. Mol. Biol. 22, 853–858 (2015).
8. A. Porro, S. Feuerhahn, P. Reichenbach, J. Lingner, Molecular dissection of telomeric repeat-containing RNA biogenesis unveils the presence of distinct and multiple regulatory pathways. Mol. Cell. Biol. 30, 4808–4817 (2010).
9. R. L. Flynn, R. C. Centore, R. J. O’Sullivan, R. Rai, A. Tse, Z. Songyang, S. Chang, J. Karlseder, L. Zou, TERRA and hnRNPA1 orchestrate an RPA-to-POT1 switch on telomeric singlestranded DNA. Nature 471, 532–536 (2011).
10. Z. Deng, Z. Wang, N. Stong, R. Plasschaert, A. Moczan, H.-S. Chen, S. Hu, P. Wikramasinghe, R. V. Davuluri, M. S. Bartolomei, H. Riethman, P. M. Lieberman, A role for CTCF and cohesin in subtelomere chromatin organization, TERRA transcription, and telomere end protection. EMBO J. 31, 4165–4178 (2012).
11. H. Episkopou, I. Draskovic, A. Van Beneden, G. Tilman, M. Mattiussi, M. Gobin, N. Arnoult, A. Londoño-Vallejo, A. Decottignies, Alternative lengthening of telomeres is characterized by reduced compaction of telomeric chromatin. Nucleic Acids Res. 42, 4391–4405 (2014).
12. R. Eid, M.-V. Demattei, H. Episkopou, C. Augé-Gouillou, A. Decottignies, N. Grandin, M. Charbonneau, Genetic inactivation of ATRX leads to a decrease in the amount of telomeric cohesin and level of telomere transcription in human glioma cells. Mol. Cell. Biol. 35, 2818–2830 (2015).
13. N. Stong, Z. Deng, R. Gupta, S. Hu, S. Paul, A. K. Weiner, E. E. Eichler, T. Graves, C. C. Fronick, L. Courtney, R. K. Wilson, P. M. Lieberman, R. V. Davuluri, H. Riethman, Subtelomeric CTCF and cohesin binding site organization using improved subtelomere assemblies and a novel annotation pipeline. Genome Res. 24, 1039–1050 (2014).
14. K. Cartharius, K. Frech, K. Grote, B. Klocke, M. Haltmeier, A. Klingenhoff, M. Frisch, M. Bayerlein, T. Werner, MatInspector and beyond: Promoter analysis based on transcription factor binding sites. Bioinformatics 21, 2933–2942 (2005).
15. R. C. Scarpulla, Nuclear control of respiratory chain expression by nuclear respiratory factors and PGC-1-related coactivator. Ann. N.Y. Acad. Sci. 1147, 321–334 (2008).
16. M. C. Chan, Z. Arany, The many roles of PGC-1a in muscle—Recent developments. Metabolism 63, 441–451 (2014).
17. S. Domcke, A. F. Bardet, P. Adrian Ginno, D. Harti, L. Burger, D. Schübeler, Competition between DNA methylation and transcription factors determines binding of NRF1. Nature 528, 575–579 (2015).
18. P. J. Fernandez-Marcos, J. Auwerx, Regulation of PGC-1a, a novel regulator of mitochondrial biogenesis. Am. J. Clin. Nutr. 93, 884S–890S (2011).
19. H. Pilegaard, B. Saltin, P.D. Neufer, Exercise induces transient transcriptional activation of the PGC-1a gene in human skeletal muscle. J. Physiol. 546, 851–858 (2003).
20. M. B. Hock, A. Kralli, Transcriptional control of mitochondrial biogenesis and function. Annu. Rev. Physiol. 71, 177–203 (2009).
21. D. C. Wright, D.-H. Han, P. M. Garcia-Roves, P. C. Geiger, T. E. Jones, J. O. Holloszy, Exerciseinduced mitochondrial biogenesis begins before the increase in muscle PGC-1a expression. J. Biol. Chem. 282, 194–199 (2007).
22. B. Viollet, L. Lantier, J. Devin-Leclerc, S. Hébrard, C. Amouyal, R. Mounier, M. Foretz, F. Andreelli, Targeting the AMPK pathway for the treatment of type 2 diabetes. Front. Biosci. 14, 3380–3400 (2009).
23. H. Izumi, R. Ohta, G. Nagatani, T. Ise, Y. Nakayama, M. Nomoto, K. Kohno, p300/CBP-associated factor (P/CAF) interacts with nuclear respiratory factor-1 to regulate the UDP-N-acetyl-a-Dgalactosamine: Polypeptide N-acetylgalactosaminyltransferase-3 gene. Biochem. J. 373, 713–722 (2003).
24. N. Gleyzer, K. Vercauteren, R. C. Scarpulla, Control of mitochondrial transcription specificity factors (TFB1M and TFB2M) by nuclear respiratory factors (NRF-1 and NRF-2) and PGC-1 family coactivators. Mol. Cell. Biol. 25, 1354–1366 (2005).
25. C. Lukas, V. Savic, S. Bekker-Jensen, C. Doil, B. Neumann, R. S. Pedersen, M. Grøfte, K. L. Chan, I. D. Hickson, J. Bartek, J. Lukas, 53BP1 nuclear bodies form around DNA lesions generated by mitotic transmission of chromosomes under replication stress. Nat. Cell Biol. 13, 243–253 (2011).
26. E. Cusanelli, C. A. Romero, P. Chartrand, Telomeric noncoding RNA TERRA is induced by telomere shortening to nucleate telomerase molecules at short telomeres. Mol. Cell 51, 780–791 (2013).
27. M. Moravec, H. Wischnewski, A. Bah, Y. Hu, N. Liu, L. Lafranchi, M. C. King, C. M. Azzalin, TERRA promotes telomerase-mediated telomere elongation in Schizosaccharomyces pombe. EMBO Rep. 17, 999–1012 (2016).
28. C. Weierich, A. Brero, S. Stein, J. von Hase, C. Cremer, T. Cremer, I. Solovei, Three-dimensional arrangements of centromeres and telomeres in nuclei of human and murine lymphocytes. Chromosome Res. 11, 485–502 (2003).
29. E. Mundstock, H. Zatti, F. M. Louzada, S. G. Oliveira, F. T. C. R. Guma, M. M. Paris, A. B. Rueda, D. G. Machado, R. T. Stein, M. H. Jones, E. E. Sarria, F. M. Barbé-Tuana, R. Mattiello, Effects of physical activity in telomere length: Systematic review and meta-analysis. Ageing Res. Rev. 22, 72–80 (2015).
30. A. Borghini, G. Giardini, A. Tonacci, F. Mastorci, A. Mercuri, S. M. Sposta, S. Moretti, M. G. Andreassi, L. Pratali, Chronic and acute effects of endurance training on telomere length. Mutagenesis 30, 711–716 (2015).
31. G. Saretzki, T. von Zglinicki, Replicative aging, telomeres, and oxidative stress. Ann. N.Y. Acad. Sci. 959, 24–29 (2002).
32. S. K. Powers, L. L. Ji, C. Leeuwenburgh, Exercise training-induced alterations in skeletal muscle antioxidant capacity: A brief review. Med. Sci. Sports Exerc. 31, 987–997 (1999).
33. E. Sahin, S. Colla, M. Liesa, J. Moslehi, F. L. Müller, M. Guo, M. Cooper, D. Kotton, A. J. Fabian, C. Walkey, R. S. Maser, G. Tonon, F. Foerster, R. Xiong, Y. A. Wang, S. A. Shukla, M. Jaskelioff, E. S. Martin, T. P. Heffernan, A. Protopopov, E. Ivanova, J. E. Mahoney, M. Kost-Alimova, S. R. Perry, R. Bronson, R. Liao, R. Mulligan, O. S. Shirihai, L. Chin, R. A. DePinho, Telomere dysfunction induces metabolic and mitochondrial compromise. Nature 470, 359–365 (2011).
34. E. Sahin, R. A. DePinho, Axis of ageing: Telomeres, p53 and mitochondria. Nat. Rev. Mol. Cell Biol. 13, 397–404 (2012).
35. A. Safdar, J. M. Bourgeois, D. I. Ogborn, J. P. Little, B. P. Hettinga, M. Akhtar, J. E. Thompson, S. Melov, N. J. Mocellin, G. C. Kujoth, T. A. Prolla, M. A. Tarnopolsky, Endurance exercise rescues progeroid aging and induces systemic mitochondrial rejuvenation in mtDNA mutator mice. Proc. Natl. Acad. Sci. U.S.A. 108, 4135–4140 (2011).
36. E. Jean, D. Laoudj-Chenivesse, C. Notarnicola, K. Rouger, N. Serratrice, A. Bonnieu, S. Gay, F. Bacou, C. Duret, G. Carnac, Aldehyde dehydrogenase activity promotes survival of human muscle precursor cells. J. Cell. Mol. Med. 15, 119–133 (2011)
37. M. Foretz, S. Hébrard, J. Leclerc, E. Zarrinpashneh, M. Soty, G. Mithieux, K. Sakamoto, F. Andreelli, B. Viollet, Metformin inhibits hepatic gluconeogenesis in mice independently of the LKB1/AMPK pathway via a decrease in hepatic energy state. J. Clin. Invest. 120, 2355–2369 (2010).
38. R. Van Thienen, G. D’Hulst, L. Deldicque, P. Hespel, Biochemical artifacts in experiments involving repeated biopsies in the same muscle. Physiol. Rep. 2, e00286 (2014).
39. J. Boros, N. Arnoult, V. Stroobant, J.-F. Collet, A. Decottignies, Polycomb repressive complex 2 and H3K27me3 cooperate with H3K9 methylation to maintain heterochromatin protein 1a at chromatin. Mol. Cell. Biol. 34, 3662–3674 (2014).
40. R. Basu, L.-T. Lai, Z. Meng, J. Wu, F. Shao, L.-F. Zhang, Using amino-labeled nucleotide probes for simultaneous single molecule RNA-DNA FISH. PLOS One 9, e107425 (2014).
41. D. J. Mahoney, K. Carey, M.-H. Fu, R. Snow, D. Cameron-Smith, G. Parise, M. A. Tarnopolsky, Real-time RT-PCR analysis of housekeeping genes in human skeletal muscle following acute exercise. Physiol. Genomics 18, 226–231 (2004).
42. E. Escoffier, A. Rezza, A. Roborel de Climens, A. Belleville, L. Gazzolo, E. Gilson, M. Duc Dodon, A balanced transcription between telomerase and the telomeric DNAbinding proteins TRF1, TRF2 and Pot1 in resting, activated, HTLV-1-transformed and Tax-expressing human T lymphocytes. Retrovirology 2, 77 (2005).
43. G. Tilman, A. Loriot, A. Van Beneden, N. Arnoult, J. A. Londoño-Vallejo, C. De Smet, A. Decottignies, Subtelomeric DNA hypomethylation is not required for telomeric sister chromatid exchanges in ALT cells. Oncogene 28, 1682–1693 (2009).
44. S. Sampl, S. Pramhas, C. Stern, M. Preusser, C. Marosi, K. Holzmann, Expression of telomeres in astrocytoma WHO grade 2 to 4: TERRA level correlates with telomere length, telomerase activity, and advanced clinical grade. Transl. Oncol. 5, 56–65 (2012).
45. N. Gonen, Y. G. Assaraf, The obligatory intestinal folate transporter PCFT (SLC46A1) is regulated by nuclear respiratory factor 1. J. Biol. Chem. 285, 33602–33613 (2010).
Acknowledgments: We thank P. Lieberman, B. Viollet, M. Foretz, K. Kohno, and G. Carnac for reagents; R. Van Thienen for muscle biopsies; all subjects for their dedication to the experiment; and N. Arnoult for critical comments on the article. Funding: A.D. has a fellowship from Télévie/ Fonds National de la Recherche Scientifique (FNRS). J.B., H.E., L.B., and A.D.C. are supported by the FNRS. F.P. has a grant from Fonds pour la formation à la Recherche dans l’Industrie et dans l’Agriculture. J.R. is supported by ProCell sprl. M.F. is supported by the Walloon Region. Author contributions: A.D. and J.B. performed most of the experiments and carried out data analysis, with assistance from F.P., J.R., M.P., and H.E. L.B. provided assistance with adenovirus production and guidance for AMPK activation. M.F. and L.D. designed and supervised the human study. A.D., L.D., and A.D.C. wrote the paper. A.D.C. conceived the project, directed the research, and assisted in data analysis. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. Correspondence and request for materials should be addressed to A.D.C. (anabelle.decottignies@uclouvain.be).
Submitted 8 January 2016 Accepted 29 June 2016 Published 27 July 2016 10.1126/sciadv.1600031
Citation: A. Diman, J. Boros, F. Poulain, J. Rodriguez, M. Purnelle, H. Episkopou, L. Bertrand, M. Francaux, L. Deldicque, A. Decottignies, Nuclear respiratory factor 1 and endurance exercise promote human telomere transcription. Sci. Adv. 2, e1600031 (2016).